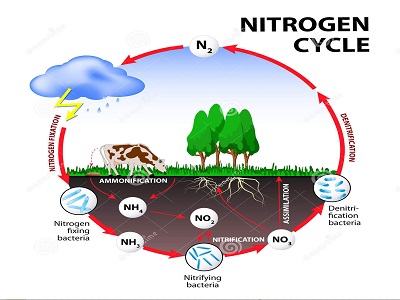
Il ciclo dell’azoto è, come dice il nome, il percorso che compie l’azoto partendo dall’atmosfera terrestre di cui è componente principale (78% contro il 21% di ossigeno), attraverso l’integrazione in sostanze organiche, per poi venire riconsegnato al suo naturale serbatoio atmosferico.
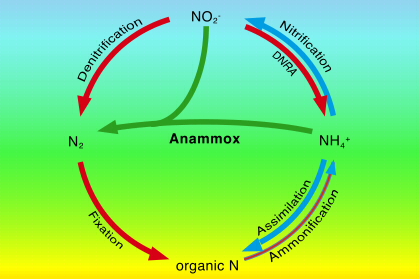
Il ciclo dell’azoto è molto complesso, anche perché l’atomo dell’azoto può entrare a far parte di un elevato numero di molecole, essenzialmente di tre tipi: Azoto molecolare (N2), Azoto organico (amminoacidi), Azoto inorganico (ammoniaca, nitriti, nitrati, sali di ammonio).
AZOTO MOLECOLARE
In chimica, la parola Azoto identifica l’elemento dal simbolo chimico N, numero atomico 7, peso atomico 14.01, raggio atomico 0.74 Å.
L’azoto molecolare invece è un gas incolore e inodore presente nell’aria. È il componente principale dell’atmosfera terrestre. La sua sigla è N2, perché a differenza dell’elemento chimico, esso è composto da 2 molecole.
È scarsamente solubile in acqua, con la quale non reagisce. È una molecola biatomica, con un triplo legame (covalente) fra i due atomi. Questo legame di molecola elementare dell’azoto con un’altra è uno dei più forti legami covalenti esistenti, ciò gli garantisce una fortissima stabilità, e per questo è detto gas inerte.
È l’elemento stabilizzante dell’atmosfera terrestre in quanto un ambiente saturo di solo ossigeno diverrebbe instabile poichè soggetto ad autocombustione.
AZOTO ORGANICO
Per azoto organico (convenzionalmente indicato con formula -NH2) si intende l’azoto presente in diversi composti organici come ammine, ammidi, immine e azoto albuminoideo. L’azoto organico rappresenta un nutriente ad azione plastica ed occupa una posizione unica tra gli elementi essenziali per la crescita degli organismi sia animali che vegetali.
AZOTO INORGANICO
L’azoto ammoniacale, nitroso e nitrico rappresentano gli ultimi stadi della decomposizione di sostanze azotate di provenienza animale e vegetale ed è il prodotto finale di scarto dei pesci.
Infatti solo i pesci producono ammoniaca come metabolita dell’azoto poiché vivendo in un mezzo acquoso ed essendo questa molto solubile, possono eliminarla senza problemi.
I processi chimici coinvolti per la formazione di composti azotati possono essere suddivisi in cinque tipi: 1) azoto fissazione; 2) immobilizzazione; 3) mineralizzazione : ammonificazione – nitrificazione; 4) denitrificazione; 5) volatilizzazione.
1) AZOTOFISSAZIONE
L’azoto come molecola non è immediatamente assimilabile dagli organismi viventi e deve quindi essere prima trasformato in sostanze chimiche che ne permettano un più facile utilizzo.
L’azoto atmosferico deve quindi prima essere attivato, cioè deve venire scisso nei due atomi di azoto che lo compongono, in modo da potersi poi legare ad altri elementi come l’idrogeno e l’ossigeno. La fase di attivazione richiede una quantità di energia pari a 160 Kcal per mole di azoto fissato.
La fissazione dell’azoto si verifica: nell’atmosfera grazie all’energia fornita dalla energia solare, dai fulmini, dai vulcani ecc. (10%); nel terreno ad opera dei microorganismi azoto fissatori (90%).
Nell’atmosfera la rottura del triplo legame dell’azoto bimolecolare produce due atomi estremamente reattivi che si legano ad ossigeno, formando ossidi di azoto, o all’idrogeno, formando ammoniaca. Queste sostanze raggiungono direttamente il suolo tramite l’acqua contenuta nelle precipitazioni.
La reazione di idrogenazione avviene nel seguente modo: N2 + 8H+ + 6e- —> 2NH4+ o anche N2 + 3H2 —> 2NH3
L’ossidazione dell’azoto avviene, invece, secondo la reazione: N2 + O2 —> 2NO
Il monossido di azoto reagisce a sua volta con l’ossigeno per dare biossido di azoto: NO + ½O2 —> NO2
Il diossido di azoto reagisce con l’acqua dell’atmosfera per dare l’acido nitrico e monossido di azoto: 3NO2 + H2O —> 2HNO3 + NO L’acido nitrico è responsabile delle cosiddette piogge acide.
Lo ione nitrato NO3– proviene dalla dissociazione completa dell’acido nitrico HNO3 o dei nitrati quando sono sciolti in acqua. Ciò avviene secondo la seguente formula: HNO3 + H2O —> H3O+ + NO3–
Nel terreno la trasformazione dell’azoto è deputata ad alcune classi di batteri.
I microorganismi fissatori di azoto si dividono in: microorganismi che vivono liberi nel terreno (fissazione non simbiontica) e microorganismi che vivono in simbiosi con le piante (fissazione simbiontica).
FISSAZIONE NON SIMBIONTICA
La reazione di fissazione dell’azoto operata dai batteri azotofissatori è la seguente: N2 + 8H+ + 8e- + 16 ATP → 2NH3 + H2 + 16ADP + 16 Pi
I gruppi più importanti di batteri che appartengono a questo gruppo sono:
batteri eterotrofi- aerobi obbligati: Azotobacter, Azotomonas, Pseudomonas, Beijerinckia;
anaerobi facoltativi: Klebsiella, Bacillus;
anaerobi obbligati: Clostridium, Methanobacillus, Desulfovibrio;
batteri fotoautotrofi, sia aerobi che anaerobi, dei generi Rhodospirillum e Rhodopseudomonas;
cianobatteri, sia aerobi che anaerobi, dei generi Nostoc, Anabaena;
funghi eterotrofi aerobi dei generi Pupillaria, Saccaromyces.
In ambienti acquatici la fissazione è operata dai cianobatteri. Questi sono organismi acquatici cosmopoliti che si possono trovare sia in acqua dolce che in acqua salata, dalle acque fredde di alta montagna alle acque termali fino a 75 °C.
Esistono sia specie planctoniche (a nuoto libero) che specie bentoniche (da fondo) o fissate al substrato. Nel mare formano pellicole nerastre sugli scogli al limite superiore dell’alta marea, e su rocce carbonatiche, dove c’è gocciolamento di acqua, formano patine lineari chiamate “strisce d’inchiostro”.
La quantità di azoto fissata dai batteri non simbionti è generalmente molto bassa in quanto limitata dalle disponibilità di energia e di carbonio, e per essere efficace questa operazione necessita di: bassi livelli di azoto presenti nel terreno; pH vicino alla neutralità; sufficiente umidità del terreno.
FISSAZIONE SIMBIONTICA
Questa fissazione è resa possibile dalla simbiosi, una stretta relazione di carattere biologico, che avviene tra microorganismi e vegetali dai quali ricevono energia e nutrimento. Qui la rottura del triplo legame covalente avviene grazie all’enzima nitrogenasi.
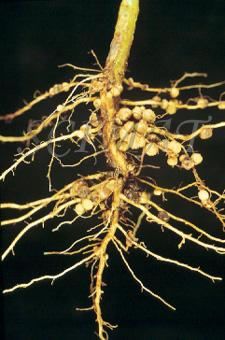
Sono azotofissatori simbionti i batteri del genere: Rhizobium e Actinomyces.
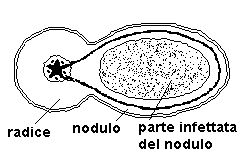
La collaborazione tra pianta e batteri avviene attraverso la produzione di azoto fissato da parte dei batteri che cedono in massima parte alla pianta, ricevendo da questa sostanze carboniose.
I batteri “infettano” la pianta penetrandola dai peli radicali, per poi trasferirsi nel tessuto corticale della radice.
La proliferazione delle cellule corticali causa la formazione del caratteristico nodulo, sede di numerose colonie batteriche.
Le condizioni ottimali per l’azione di questi batteri sono: pH vicino alla neutralità; buona aerazione del terreno; disponibilità di fosforo; temperatura elevata del terreno; presenza adeguata di metalli pesanti nel terreno (quali molibdeno, rame, manganese, cobalto, ferro, ecc.) che agiscono da catalizzatori per le reazioni.
 |  |
2) IMMOBILIZZAZIONE
È definita come la trasformazione dell’azoto minerale in azoto organico. I batteri assumono azoto minerale (inorganico) per incorporarlo nella loro biomassa. L’azoto viene sottratto all’utilizzo da parte delle piante.
3) MINERALIZZAZIONE
La mineralizzazione dell’azoto è costituita da una serie di reazioni attraverso le quali l’azoto, contenuto nella sostanza organica, viene convertito in azoto inorganico. Inizialmente in forma ammoniacale e poi in quella nitrica.
Durante questo processo vengono liberati, in forma minerale, gli elementi nutritivi e l’azoto, per essere resi disponibili all’assorbimento da parte delle piante.

3a) AMMONIFICAZIONE
La prima tappa della complessa serie di reazioni che compongono la mineralizzazione viene definita ammonificazione.
Essa consiste nella trasformazione di sostanze organiche contenenti azoto, sotto forma di idrati di ammonio. L’ammonificazione ha luogo sia nel terreno che nell’acqua. In entrambe le circostanze il processo di ammonificazione è il risultato della ripartizione della materia organica, come animali morti e piante o materiali di scarto come gli escrementi.
Queste sostanze vengono decomposti da: batteri appartenenti ai generi Bacillus, Clostridium, Pseudomonas; attinomiceti del genere Streptomyces; funghi dei generi Aspergillus, Pennicillium, Fusarium.
La degradazione delle proteine porta alla formazione di ammoniaca ed ha inizio con la scissione delle proteine in aminoacidi da parte degli enzimi proteolitici (proteasi e peptidasi) emessi dai microrganismi decompositori.
Questa è la sequenza delle reazioni della proteolisi, o idrolisi enzimatica delle proteine:
proteasi peptidasi
PROTEINE ——————–> PEPTIDI ——————–> AMINOACIDI
Gli aminoacidi formati con la proteolisi subiscono a loro volta una degradazione microbica detta di deaminazione ossidativa, che consiste in una rimozione del gruppo amminico (NH2). Il prodotto finale è l’ammoniaca, mentre gli atomi di carbonio vengono convertiti in glucosio.
L’ammoniaca (NH3) è un composto volatile e può abbandonare il suolo o l’acqua che la contiene in soluzione (volatilizzazione), ma se viene solubilizzata dà origine allo ione ammonio (NH4+), secondo la seguente reazione: NH4+ —> NH3 + H+
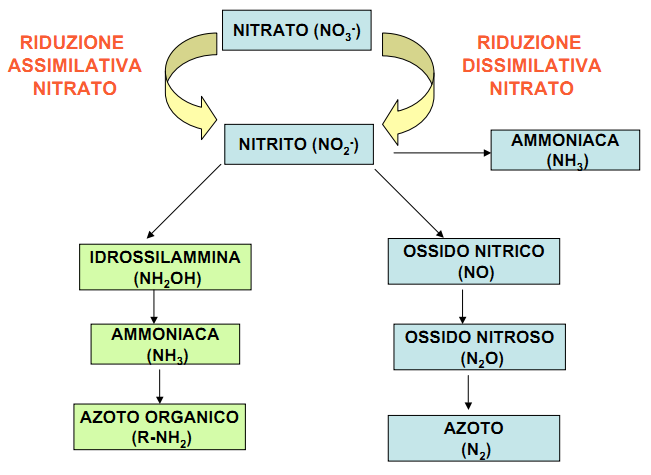
3b) NITRIFICAZIONE
Le tappe successive della mineralizzazione prendono il nome di nitrificazione e consistono nella progressiva ossidazione dell’ammonio in nitriti, e poi nitrati, nei processi che prendono il nome di nitritazione e nitratazione, operati da alcuni tipi di batteri chemioautotrofi (Nitrosomonas e Nitrobacter principalmente).
Entrambi i processi, per realizzarsi in modo ottimale, richiedono: elevati tassi di ossigeno (saturazione superiore all’80%); specifici e limitati intervalli ottimali di pH (Nitrosomonas 7,8-8,0 – Nitrobacter 7,3-7,5); temperature ottimali da 25°C a 30°C; degradano l’attività rapidamente sotto i 10-12°C; che le reazioni avvengano in assenza di luce.
I batteri responsabili di queste trasformazioni presentano inoltre: basso tasso di riproduzione (24-36 ore); non sono sporigeni e solo i Nitrosomonas, in assenza di nutrimento, entrano in uno stato di dormienza; muoiono rapidamente in assenza di cibo; sono sensibili a shock osmotici; in acqua devono aderire ad un substrato per esercitare il loro compito ossidativo.
NITRATAZIONE
Durante la nitritazione lo ione ammonio viene trasformato in nitrito ad opera di una classe di batteri denominati batteri nitrosi che sono: Nitrosomonas, Nitrosococcus, Nitrosospira.
La reazione chimica è la seguente: NH4+ + 1,5O2 —> NH2OH + H
La reazione trasforma l’ammoniaca dapprima in idrossilamina ed è catalizzata dall’enzima ammonio mono-ossigenasi. Poi l’idrossilamina viene trasformata in nitrito secondo una reazione catalizzata dall’enzima idrossilamina ossidoreduttasi: NH2OH + O2 —> H+ + H2O + NO2−
Come si può osservare la sequenza di queste reazioni consuma alcalinità.
Questo valore quindi, nell’acqua, deve essere necessariamente mantenuto sufficiente a garantire un suo potere tampone tale da evitare oscillazioni del pH.
NITRATAZIONE
Durante la nitratazione le molecole del nitrito vengono convertite in nitrato ad opera dei batteri nitrici: Nitrobacter, Nitrospira, Nitrococcus, secondo la reazione, catalizzata dall’enzima nitrito ossidoreduttasi: NO2− + 0,5O2 —> NO3−
Questi batteri non utilizzano tutta l’ammoniaca come fonte di energia. Parte di questa infatti viene dirottata per la sintesi cellulare. Considerando questo fatto, la reazione complessiva della nitrificazione risulta essere la seguente: NH4+ + 1,863 O2 + 0,098 CO2 —> 0,0916 C5H7NO2 + 0,98 NO3– + 1,98 H+ + 1,98 H2O
Mentre per il processo di formazione di nuove cellule la reazione sarà: NH4+ + 2HCO3– + 2O2 —> NO2– + 2CO2 + H2O
4) DENITRIFICAZIONE
La denitrificazione consiste nella riduzione dei nitrati e dei nitriti in composti gassosi come l’azoto bimolecolare (N2) o il protossido di azoto (N2O).
Diversamente da quanto accade per la nitrificazione, numerosi sono i ceppi di batteri che possono dare luogo alla denitrificazione. Questi organismi capaci di utilizzare nitrati e/o nitriti per la respirazione cellulare, sono in gran parte eterotrofi e facoltativi.
Sono eterotrofi in quanto necessitano, per il loro metabolismo, di una fonte organica di carbonio, ad esempio gli zuccheri, e facoltativi perchè possono utilizzare, come accettore di elettroni, indifferentemente sia l’ossigeno che l’azoto nitrico/nitroso.
In ogni caso la compresenza di ossigeno e nitrati vede il consumo preferenziale dell’ossigeno perchè energeticamente più favorevole al metabolismo batterico.
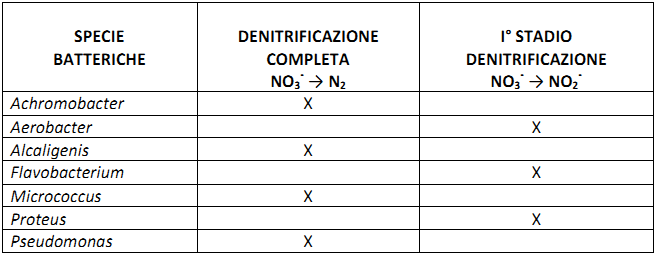
Il processo di denitrificazione è attuato da diversi batteri, dei quali, alcuni partecipano a tutto il processo, ed altri solo a determinate fasi. Essi possono essere suddivisi in tre gruppi:
1) Batteri che possono solo ridurre il nitrato NO3– a nitrito NO2– ;
2) Batteri che possono ridurre il nitrito ma non il nitrato poichè sono mancanti dell’enzima nitrato reduttasi;
3) Batteri che riducono il nitrato ad ossido nitroso N2O secondo tre vie: in anaerobiosi per riduzione dei nitrati; in anaerobiosi durante l’assimilazione del nitrato; in anaerobiosi durante l’ossidazione di ammoniaca NH4+ a nitrato NO3–
Esistono due differenti tipi di denitrificazione:
DENITRIFICAZIONE ASSIMILATIVA
(avviene nei vegetali superiori, alghe, batteri, ed alcuni funghi): l’NH4+ viene ottenuto, in condizioni aerobiche, dall’uso del nitrato come fonte di azoto per la sintesi delle proteine. Non causa una perdita di azoto, ma una temporanea immobilizzazione;
DENITRIFICAZIONE DISSIMILATIVA
detta anche denitrificazione vera. Il nitrato NO3– viene usato come accettore di elettroni in reazioni di ossidazione. È la vera denitrificazione, dove il nitrato sostituisce l’O2 in un meccanismo respiratorio.
Il processo è dovuto all’azione di alcuni batteri eterotrofi facoltativi anaerobi dei generi: Pseudomonas, Bacillus, Spirillum, Hyphomicrobium, Agrobacterium, Acinetobacter, Propionobacterium, Rhizobium, Cornebacterium, Cytophata, Thiobacillus,Moraxella, Neisseria, Vibrio, Flavobacterium e Alcaligenes.
Questi sono batteri che in condizioni di mancanza di ossigeno atmosferico utilizzano per la loro respirazione l’ossigeno contenuto nei nitrati.
Nitrati così ridotti a protossido di azoto ed azoto elementare che, essendo per loro natura dei gas, si liberano nell’atmosfera.
I batteri denitrificanti possono essere autotrofi oppure eterotrofi.
I primi comprendono Micrococcus denitrificans (autotrofo facoltativo) e Thiobacillus denitrificans. I secondi comprendono numerosi generi tra i quali Micrococcus, Pseudomonas, Achromobacter e Bacillus.
La denitrificazione ha luogo secondo quattro stadi di trasformazione chimica catalizzati da differenti enzimi:
stadio 1 – NO3− +2e− +2H+ —> NO2− + H2O
stadio 2 – 2NO2- + 4H+ + 2e− —> 2NO2− + 2H20
stadio 3 – 2NO + 2H+ + 2e− —> N2O + H20
stadio 4 – N2O + 2H+ + 2e− —> N2 + H20
La reazione di denitrificazione complessiva (esclusa la degradazione delle molecole organiche eventualmente associata) è la seguente: 2NO3– + 12H+ + 10e- —> N2 + 6H20
A differenza delle reazioni di nitrificazione, la denitrificazione nel suo complesso è un processo che restituisce alcalinità all’acqua (o al terreno) dalla quale è stata sottratta durante le precedenti reazioni ossidative dell’ammonio.
I batteri denitrificanti (facoltativi anaerobi) svolgono la loro attività: a intervalli di pH compresi tra 7,0 e 9,0; in quasi totale assenza di ossigeno; senza essere influenzati dalla luce solare; a temperature dai 20 ai 33°C; sotto i 5°C muoiono; con tempi di riproduzione di 15-30 minuti; sono sporigeni ed ubiquitari; sono scarsamente sensibili agli shock osmotici; in acqua non devono aderire ad un substrato per essere attivi metabolicamente.
ALTRE FORME DI NITRIFICAZIONE E DENITRIFICAZIONE
Esistono poi altre forme di nitrificazione/denitrificazione di cui si è venuti solo recentemente a conoscenza: AOA (Ammonia-Oxidizing Archaea); DNRA (Dissimilatory Nitrate/nitrite Reduction to Ammonium); ANAMMOX (ANaerobic AMMonium OXidation)
AOA – Archeobatteri che ossidano l’ammonio (nitrificanti)
Il nome di questi microrganismi fa riferimento alla loro antica origine. Questi batteri sono in grado di vivere in ambienti estremi dove la sopravvivenza è difficile per molte forme di vita. L’ambiente in cui vivono è probabilmente molto simile a quello presente sulla terra durante le prime fasi di sviluppo dei procarioti.
Sono costituiti da singole cellule mancanti di nucleo e assieme ai batteri sono stati classificati come procarioti. Tuttavia questa classificazione è stata più volte rimaneggiata ed è tutt’ora oggetto di studio.
Esistono sottogruppi di questi batteri che vivono in condizioni di estrema salinità dell’acqua (alofili), oppure in habitat molto acidi e molto caldi (60-80°C e pH 2-4) e vengono denominati termoacidofili, oppure vi sono i metanogeni che producono metano in condizioni di totale mancanza di ossigeno.
Nonostante gli ambienti estremi in cui riescono a vivere, gli archeobatteri sono presenti anche in ambienti più ospitali.Gli archeobatteri si distinguono quindi dagli eubatteri che sono invece definiti come un sottoregno dei bacteria (ovvero i batteri).
Le profonde differenze evolutive che dividono gli eubatteri dagli archeobatteri non erano evidenti sulla base del fenotipo, mentre divenne chiaro solo dopo il confronto delle loro sequenze nucleotidiche. L’ipotesi che esistano tre gruppi evolutivi principali in seno agli organismi è confortata anche dalle sequenze di altri geni come quelle dei geni per alcune proteine di base, per alcuni RNA ribosomiali. Questi tre gruppi sono Crenarchaeota, Euryarchaeota e Korarchaeota.
Gli archea sono in grado di: ossidare l’ammonio a nitrito senza che vi sia il normale passaggio attraverso l’idrossilamina; possono sopportare la disidratazione, tanto da vivere in acque satura di sale (Halobacterium); sono stati trovati sui fondali oceanici vicino a bocche eruttive a temperature di 120° ed oltre e a pressioni elevatissime che impediscono l’ebollizione dell’acqua; la loro parete è priva di peptidoglicani, catene lineari di polisaccaridi che costituiscono le pareti cellulari degli Eubatteri; le loro membrane cellulari sono costituite da catene di isoprene e non da fosfolipidi; possiedono un pigmento sensibile alla luce rossa, la alorodopsina (molto simile alla rodopsina, presente anche nella retina dei Vertebrati) che serve loro per ottenere energia.
DNRA – Riduzione anaerobica dei nitrati ad ammonio (denitrificanti)
Molto spesso ci si riferisce al processo di denitrificazione come ad una sequenza di reazioni che, partendo dai nitrati, passando dai nitriti ed arrivando ad ammonio porti successivamente alla liberazione di azoto gassoso: NO3– —> NO2– —> NH4+ —> N2
Questa ovviamente è una semplificazione, o meglio una sintesi di due importanti meccanismi di denitrificazione che si servono di distinti batteri ed enzimi per arrivare a quell’obiettivo: la denitrificazione vera e la DNRA.
Denitrificazione vera (dissimilativa):
NO3− —> NO2− —> 2NO2– —> N2O —> N2DNRA
NO3− —> NO2− —> NH4+
Queste avvengono in condizioni simili di basse concentrazioni di ossigeno, ma mentre la denitrificazione dissimilativa trasforma il nitrato in nitrito, da qui in ossido nitrico e passando per l’ossido nitroso, arriva ad azoto gassoso, senza passare per l’ammoniaca, nella DNRA il passaggio da nitrato prevede dapprima la trasformazione in nitrito e poi subito in ammoniaca. Qui la reazione mediata da questi batteri si interrompe. Questo processo viene anche definito ammonificazione fermentativa.
 L’importanza della DNRA sta nel fatto che il nitrato viene convertito in una diversa forma minerale di azoto che è meno mobile e, sebbene sia in contrasto con la denitrificazione, essa è in grado di conservare l’azoto nell’ecosistema senza disperderlo nell’atmosfera sotto forma di gas.
L’importanza della DNRA sta nel fatto che il nitrato viene convertito in una diversa forma minerale di azoto che è meno mobile e, sebbene sia in contrasto con la denitrificazione, essa è in grado di conservare l’azoto nell’ecosistema senza disperderlo nell’atmosfera sotto forma di gas.
Per questo motivo la DNRA è stata definita come un cortocircuito nel ciclo biologico dell’azoto, in quanto il diretto trasferimento del nitrato in nitrito e poi ammoniaca bypassa la denitrificazione e la fissazione.
Le due reazioni della DNRA sono catalizzate da due enzimi di cui uno, il primo (da nitrato a nitrito) è respiratorio, mentre il secondo (da nitrito ad ammoniaca) è fermentativo.
La DNRA è stata ampiamente studiata in contesti acquatici (in particolare negli ecosistemi marini), mentre non ci sono ancora studi approfonditi su questo processo quando avviene nel suolo.
La presenza delle radici nel suolo modifica la denitrificazione come conseguenza dell’alterazione del substrato e della disponibilità di ossigeno ed è quindi ormai assodato che la denitrificazione è stimolata dalla presenza di radici. Ma questi dati sono disponibili solo per ambienti umidi e per piante acquatiche.
Uno studio in tal senso ha evidenziato come in un vaso piantumato l’attività della DNRA fosse del 53% contro un 71% a vantaggio della denitrificazione che si verifica in un terreno non piantumato. Tuttavia il contributo delle radici al processo DNRA nel suolo si è attestato essere solo dell’1% in più rispetto allo stesso terreno non piantumato.
Un altro studio condotto su zone acquitrinose in periodo di riposo vegetale, la DNRA predominava sulla denitrificazione.
Una ritenzione di azoto significativa è stato ottenuta in uno studio in cui la DNRA avveniva in terreni piantumati di suoli montagnosi tropicali.Oltre l’assimilazione da parte della pianta, la ritenzione di azoto nel terreno può essere dovuta all’assorbimento di ammoniaca prodotta tramite processo DNRA su terreni argillosi (in ragione della opposta carica elettrica) o sostanze organiche.
Sebbene il processo DNRA non preveda l’intermedio dell’ossido nitroso (N2O) tipico della denitrificazione, sono stati scoperti ceppi batterici (non ancora identificati precisamente) capaci di tale reazione. Gli studiosi suggeriscono che questo sistema rappresenti un meccanismo di detossificazione per evitare alte concentrazioni di nitrito.
L’impegno detossificante in un batterio con metabolismo DNRA non supera comunque l’1% della riduzione da nitrito a nitrato.Da studi condotti sono stati evidenziati diversi generi di batteri capaci di processi DNRA:
– Clostridium
– Citrobacter
– Enterobacter
– Erwinia
– Escherichia
– Klebsiella
– Bacillus
– Pseudomonas
– Arthrobacter
ANAMMOX – Ossidazione anaerobica dell’ammonio (denitrificanti)
La reazione anammox è importante in quanto fino a qualche anno fa non si erano ancora individuati ceppi batterici capaci di metabolizzare l’ammonio in condizioni anaerobiche. Pertanto si credeva che l’ammonio nei sedimenti dei bacini idrici giacesse immodificato ed inerte.
Nel 2008 vennero isolati per la prima volta dei batteri marini appartenenti al genere Scalindua. Dopo di allora molti altri sono stati individuati ed appartengono all’ordine dei Planctomycetesdi tra cui Planctomyces e Pirellula sono i generi più noti.
Altri generi sono al vaglio: Brocadia, Kuenenia, Anammoxoglobus, Jettenia (tutti di acqua dolce) e Scalindua (di acqua marina).
L’ammonio è ossidato ad azoto molecolare usando il nitrito come accettore di elettroni. La reazione è la seguente: NH4+ + NO2– –> N2 + 2H2O
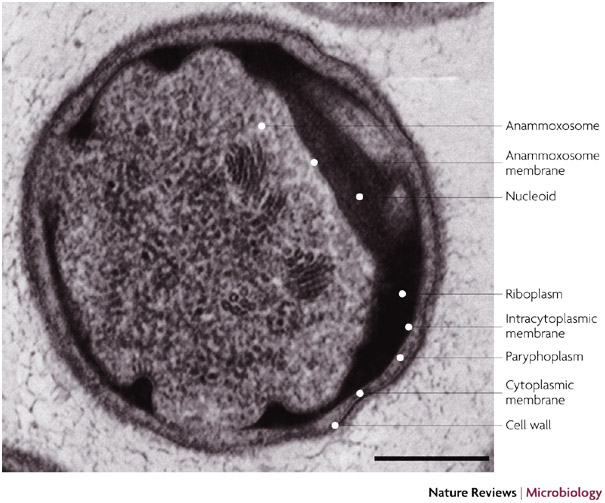
Questa reazione batterica avviene nell’anammoxosoma che è un compartimento intracitoplasmatico racchiuso da una membrana a singolo stato lipidico.
Tale struttura appare inusuale, dato che rappresenta un organello che è più tipicamente presente negli organismi eucarioti.
Il processo Anammox è strettamente autotrofo e anaerobico e prevede l’idrossilamina come intermedio di passaggio. L’ammonio con idrossilamina è convertito ad idrazina e successivamente questa è ossidata ad azoto bimolecolare.
La presenza della membrana dell’anammoxosoma ha anche il significato di isolare dal resto della cellula l’idrazina in quanto sostanza altamente reattiva.
L’importanza delle reazioni anammox risultano evidenti quando accostate a quelle del processo DNRA sopradescritto, infatti se poste in sequenza DNRA —> Anammox si realizza una completa ossidazione da nitrato ad azoto molecolare in condizioni del tutto anaerobiche.
Da studi condotti pare che il contributo complessivo della reazione anammox negli oceani alla produzione di azoto gassoso sia del 50% nei confronti di tutto l’azoto prodotto e liberato nell’atmosfera.
5) VOLATILIZZAZIONE
Perdite di azoto ammoniacale si possono verificare per evaporazione in una soluzione in cui vi sia dell’ammoniaca. Vediamo come.
Gli ioni ammonio (NH4+) sono principalmente molecole di ammoniaca anidra (NH3+) a cui legato uno ione idrogeno (H+). Quando uno ione idrogeno viene rimosso da una molecola di ammonio da un altro ione idrossile (OH–), la molecola risultante di ammoniaca può evaporare o volatilizzarsi.
Questo fenomeno è maggiormente presente in soluzioni ad alto pH che contengano grandi quantità di ioni idrossile (OH–).
Perchè nel terreno la volatilizzazione possa avere luogo è necessario che siano presenti ioni ammonio in soluzione, in quanto la reazione non si applica agli ioni ammonio adsorbiti dalla capacità di scambio cationico del terreno (per effetto dell’attrazione di cariche opposte).
La velocità della volatilizzazione dell’ammoniaca dipende dalla temperatura e dalle condizioni del vento, all’aumentare delle quali l’equilibrio tra ammonio e ammoniaca (NH4+ <—> NH3 + H+) si sposta verso la formazione di ammoniaca (a destra nella formula), provocando un aumento della costante di volatilizzazione.
RIDUZIONE ASSIMILATIVA DELL’AZOTO NEI VEGETALI (O IMMOBILIZZAZIONE)
Per le piante l’azoto, tra tutti gli elementi, è quello maggiormente importante per lo sviluppo.
È un componente che entra a far parte di numerosi composti fra cui le proteine, gli enzimi, gli acidi nucleici, la clorofilla e alcune vitamine, tutti essenziali per lo svolgimento delle funzioni biologiche. Una sua carenza provoca la riduzione della crescita della pianta, con graduale clorosi delle foglie e conseguente accumulo di carboidrati che inducono la sintesi delle antocianine nelle foglie e negli steli della pianta.
Una volta assorbito, l’azoto è in parte utilizzato nelle radici, ed in parte trasportato al fusto, alle foglie ed ai semi. Anche un eccesso di azoto provoca problemi alla pianta, tra i quali rallentamento del suo sviluppo, aumento del consumo idrico, maggior suscettibilità alle variazioni climatiche e alle infestazioni parassitarie.
Le piante sono in grado di estrarre dal terreno, o da una soluzione di acqua, lo ione ammonio e lo ione nitrato. Sebbene nel terreno la concentrazione di ammonio sia grandemente inferiore rispetto a quella del nitrato, questa differenza non si riflette necessariamente in una diversa utilizzazione delle due sorgenti da parte della pianta.
Una ragione fondamentale di questo comportamento sta nel fatto che l’utilizzo di ammonio da parte della pianta richiede meno energia per l’assimilazione rispetto al nitrato il quale, per poter essere assimilato dalla pianta, deve venire prima ridotto in nitrito e poi in ammonio da parte di due enzimi della pianta, la nitrato reduttasi e la nitrito reduttasi che catalizzano le seguenti reazioni a partire da acido nitrico (HNO3) e acido nitroso (HNO2) rispettivamente: HNO3 + 2e + 2H+ ——–> HNO2 + H2O nitrato reduttasi
La riduzione da nitrito ad ammoniaca avviene secondo la reazione: HNO2 + 6e + H+ ——–> NH3 + H2O nitrito reduttasi
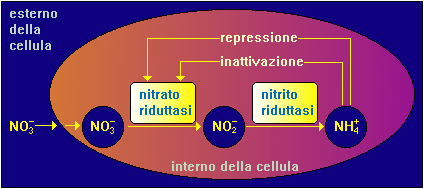
Un altro motivo che favorisce l’assorbimento di nitrato è rappresentato dal fatto che nei terreni la carica positiva, rappresentata dagli ioni ammonio, è attratta dalla carica negativa presente nell’argilla del terreno, la quale rende difficile l’assorbimento dell’ammonio nella radice della pianta.
Lo ione nitrato viene assorbito dalla pianta attraverso due diversi meccanismi.
Il primo, detto penetrazione passiva, avviene quando la concentrazione di nitrati della soluzione circolante del terreno supera quella dello spazio libero della radice.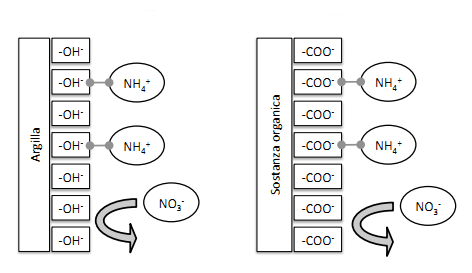
Il secondo, detto penetrazione attiva, avviene quando la concentrazione di nitrati all’interno delle cellule radicali è superiore a quella esterna. Quest’ultimo sistema richiede un sistema specifico di trasporto fornito dalle proteine attraverso un consumo di energia.
Lo ione nitrato invece essendo carico negativamente si trova quasi completamente disciolto nella soluzione circolante del terreno all’interno del quale si muove, per flusso di massa, e può raggiungere con estrema facilità la superficie della radice e venire quindi assorbito. In un sistema acquatico, invece, lo ione ammonio rimane in soluzione come quello del nitrato, quindi la pianta, dovendo operare una scelta di convenienza energetica, si orienta sull’ammonio.
ORGANICAZIONE
Una volta presente all’interno della pianta, l’ammonio deve venire rapidamente convertito in amminoacidi (organicazione), dato che elevate concentrazioni risultano tossiche per la stessa.
La tossicità per l’ammonio può essere dovuta al fatto che il suo trasporto è legato al rilascio di protoni (H+) che provocano una diminuzione del pH con acidificazione dell’ambiente intracellulare. L’assimilazione dell’ammonio avviene principalmente ad opera del sistema enzimatico della glutamina sintetasi e dellaglutamato sintasi. Gli amminoacidi ottenuti da queste reazioni vengono successivamente trasportati nei vari tessuti ed organi della pianta ad opera dei vasi xilematici.
In alcune condizioni di crescita l’azoto può essere anche assimilato nelle foglie e da queste, in forma principalmente di amminoacidi, viene trasferito alle radici attraverso i vasi floematici.
Un interessante approfondimento di questo argomento lo trovate in questo articolo: Nitrogen Metabolism

Le seguenti reazioni chimiche rappresentano il destino del nitrato nella pianta durante il giorno e durante la notte:
HNO3 + H2O —> NH3 + 2O2 (di giorno)
2HNO3 + C6H12O6 + 2O2 —> 2NH3 + 4H2O + 6CO2 (di notte)
NITROSOMONAS E NITROBACTER
Le specie del genere Nitrosomonas appartengono alla famiglia delle Nitrobacteraceae che raggruppa due gruppi di batteri Gram negativi responsabili del processo di nitrificazione.
Il primo gruppo ossida l’ammoniaca a nitrito (Nitrosomonas), il secondo ossida il nitrito a nitrato (Nitrobacter).
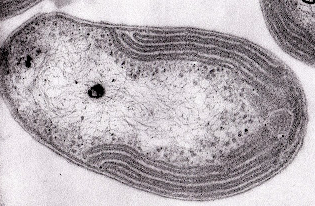
I Nitrosomonas sono batteri aerobi obbligati chemioautotrofi a forma di bastoncello. Possono essere mobili o immobili a seconda del genere. Sono in grado di generare una complessa serie di membrane interne che formano dei tubi lunghi e stretti all’interno della cellula.
Questi sono originati dalla membrana cellulare con la quale rimangono in contatto. In queste membrane avviene la reazione di ossidazione dell’ammonio a nitrito.
Questi batteri ottengono il carbonio dall’atmosfera tramite il processo di fissazione del carbonio che trasforma il carbonio inorganico (anidride carbonica) in carbonio organico. Per ottenere l’energia necessaria a questa trasformazione, i Nitrosomonas convertono l’ammonio in nitrito, a differenza delle piante che si servono dell’energia solare per mezzo della fotosintesi.
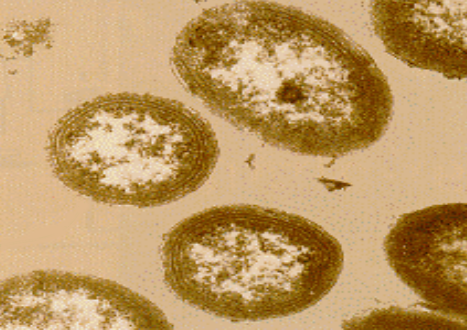
Sono molto sensibili alla luce e tendono a riprodursi in anfratti lontani dalla luce diretta del sole.
Questi batteri colonizzano il suolo, le acque dolci e marine e appaiono in coppia o in corte catene, spesso si osservano forme cocciche (sferiche).
I Nitrobacter sono generi di batteri specializzati nella trasformazione di nitrito in nitrato. Sono chemioautotrofi, ovvero organismi autotrofi che sfruttano l’energia derivante da reazioni chimiche di ossidazione tra sostanze inorganiche per produrre materia organica.
Il basso rendimento energetico della loro reazione di ossidazione dell’ammonio rende la loro capacità di riproduzione molto lenta e può durare quindi anche svariati giorni.
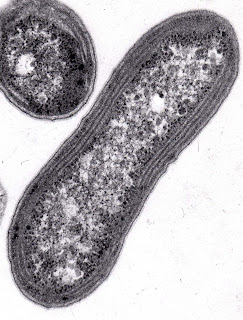
Possono avere una forma a bastoncello, a pera o essere pleomorfi (organismi privi di una forma stabile e definita). Sono dotati di un flagello.
Presentano delle inclusioni citoplasmatiche chiamate carboxisomi e granuli di PHB (acido poli-b-idrossibutirrico), un composto di tipo lipidico formato da unità ripetute di acido beta-idrossibutirrico.
Queste inclusioni hanno un significato di riserva energetica per il batterio. I Nitrobacter colonizzano il suolo, le acque dolci e marine.
Le specie nel genere comprendono Nitrobacter winogradskyi, Nitrobacter hamburgensis, Nitrobacter vulgaris e Nitrobacter alkalicus.
NITROCOCCUS
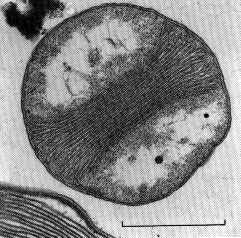
Originariamente tutti i batteri nitrificanti erano classificati sotto la famiglia delle Nitrobacteraceae, poi un sequenzamento del DNA ha evidenziato come questo batterio rappresenti una specie molto diversa, appartenente alla famiglia delle Ectothiorhodospiraceae.
Il Nitrococco mobilis si trova unicamente in acque marine poichè è un alofilico obbligato (che ama il sale). Ha una forma sferica (unico tra i batteri nitrificanti) ed un flagello per aiutarsi nel movimento.
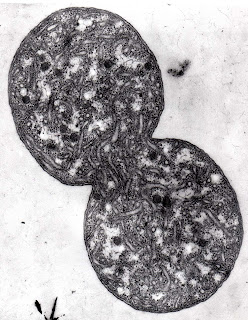
È un batterio nitrificante molto importante nelle acque marine ed è un chemioautotrofo obbligato, l’energia necessaria al suo metabolismo proviene unicamente dalla reazione chimica di ossidazione del nitrito a nitrato.
La maggior quota di questa energia (80%) viene impiegata per la fissazione del carbonio. Ne deriva, come tutti gli altri batteri nitrificanti, una lenta capacità di replicazione.
La carenza di ossigeno non danneggia il batterio, ma ne deprime ulteriormente la riproduzione.
È dotato di un sistema di microtubuli prodotti dalla invaginazione della membrana plasmatica nei quali è contenuto l’enzima nitrito ossidoreduttasi responsabile della conversione del nitrito in nitrato.
Questi microtubuli hanno una disposizione intracellulare tipicamente disordinata in questo batterio.
Nel citoplasma sono presenti dei corpi inclusi costituiti da aggregati di proteine insolubili. Sono inoltre rinvenibili depositi di glicogeno e granuli di PHB.
NITROSOSPIRA
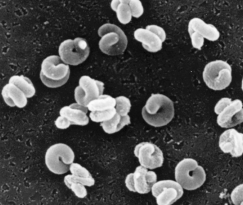
La Nitrosospira appartiene alla famiglia delle Nitrosomonadaceae ed è un batterio Gram-negativo chemiolitoautotrofo che ossida ammonio a nitrito come sorgente di energia, ed assorbe CO2 come fonte preferenziale di carbonio.
Possiede una forma vibrioide (a virgola o a “C”) molto caratteristica.
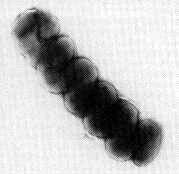
La Nitrosospira è maggiormente suscettibile alle variazioni di pH rispetto ai Nitrosomonas per le quali oltre un valore di 8,0-8,5 questi batteri muoiono.
È diffuso nel suolo dove conduce vita libera. Non è dotato di flagello pertanto è immobile.
Possiede tempi di riproduzione superiori ai Nitrobacter, e studi condotti sulle colonie batteriche degli acquari hanno evidenziato una preponderante presenza di questo batterio rispetto al Nitrobacter.
Questo spiegherebbe la relativa inefficacia di alcuni prodotti di integrazione batterica usati in acquariologia che contengono unicamente ceppi di Nitrobacter.
NITROSOCOCCUS
Il Nitrosococcus oceani è un batterio nitrificante Gram- chemiolitoautotrofo appartenente alla famiglia delle Chromatiaceae.
La maggior parte della sua energia deriva dalla ossidazione dell’ammonio in nitrito, tuttavia partecipa anche al ciclo dello Zolfo, dato che riduce il solfato in acido solfidrico (H2S).
È il primo batterio nitrificante isolato dalle acque oceaniche.
Al tempo era noto come Nitrosocystis oceanus.
Il Nitrococcus oceani è in grado di formare delle cisti ed assumere un aspetto gelatinoso attraverso la secrezione di sostanze polisaccaridiche e composti dell’azoto in situazioni di difficoltà.
È interessante notare come il Nitrosococco presenti entrambe le funzioni nitrificanti e denitrificanti.
Possiede una forma sferica o ellissoidale, ed un flagello posto in zona polare usato per il movimento.
Nel citoplasma si evidenzia la presenza di un reticolo di microtubuli che attraversa nel mezzo la cellula similmente ad altri batteri ammonio ossidanti.
CLASSIFICAZIONE DEGLI ORGANISMI IN BASE ALLE FONTI NUTRITIVE
• AUTOTROFI: organismo in grado di biosintetizzare il materiale cellulare utilizzando CO2 come unica fonte di carbonio.
• ETEROTROFI: organismi che utilizzano substrati organici preformati come fonte principale di carbonio.
• CHEMIOTROFI: ricavano energia dall’ossidazione di composti chimici organici o inorganici.
• FOTOAUTOTROFI: i fotoautotrofi utilizzano la luce come fonte di energia e CO2 come fonte principale di carbonio. Per fotoautotrofo obbligato si intende un organismo che cresce solo in presenza di luce e CO2; inoltre usa sostanze inorganiche quali H2O, H2, o H2S, come donatori di elettroni per ridurre CO2 in carbonio cellulare (CH2O).
• FOTOETEROTROFI: crescono mediante fotosintesi se dotati di un donatore di elettroni (H2 o organico) per l’assimilazione riduttiva di CO2. Questi microrganismi possono inoltre utilizzare la luce come fonte di energia e contemporaneamente assimilare dall’ambiente composti organici come substrato per la crescita.
• CHEMIOAUTOTROFI: i chemioautotrofi utilizzano substrati inorganici ridotti sia per l’assimilazione riduttiva di CO2 sia come fonte di energia; questi microrganismi ricavano l’energia necessaria dalla ossidazione di H2, NH3, NO2, H2S o Fe2+. I chemioautotrofi aerobici utilizzano O2 come accettore terminale di elettroni, mentre alcuni Archaea anaerobici usano lo zolfo inorganico.
• CHEMIOETEROTROFO: sono microrganismi che assimilano le sostanze organiche come fonte sia di carbonio che di energia; tale fonte può essere rappresentata da un unico substrato (glucosio) oppure vi possono essere fonti differenti di carbonio e di energia. Per esempio i riduttori del solfato utilizzano H2 per ottenere energia ma necessitano di un substrato organico per la biosintesi cellulare.
CLASSIFICAZIONE DEI BATTERI IN BASE ALLA MODALITA’ DI RESPIRAZIONE
Dalla trattazione finora fatta emergono quattro tipologie di batteri, ognuna con le seguenti caratteristiche:
Batteri Aerobi Obbligati: quelli responsabili della nitrificazione (Nitrosomonas, Nitrobacter, ecc.)
Questi sono organismi in grado di sopravvivere in presenza di ossigeno che viene impiegato come accettore di elettroni nelle reazioni di ossidazione dell’azoto inorganico. Questi batteri dovono essere in grado di contrastare obbligatoriamente alti livelli di stress ossidativo. Per questo motivo i batteri aerobi obbligati contengono nel citoplasma enzimi antiossidanti quali la superossido dismutasi, perossidasi e catalasi.
Batteri Aerobi Facoltativi: rappresentati dai generi Stafilococchi Streptococchi, Entrococchi, Klebsiella
I batteri aerobi facoltativi possono utilizzare l’ossigeno, oppure altre molecole, in stato ossidato. Questi organismi sono quindi in grado di sopravvivere in assenza di ossigeno. Rimane tuttavia che il loro metabolismo principale sia quello aerobico, ed in questo si differenziano dai batteri anaerobi facoltativi, il cui metabolismo principale è quello anaerobico. Fanno parte di questo gruppo principalmente alcuni generi di batteri di interesse medico.
Batteri Anaerobi Obbligati: Clostridium, batteri solfato-riduttori
Questi batteri non sono in grado di vivere in presenza di ossigeno in quanto non sono dotati di enzimi ad azione antiossidante, in grado di proteggerli dallo stress ossidativo generato dall’ossigeno.
Tra questi batteri vi sono quelli solfato-riduttori che una volta venivano considerati a metabolismo strettamente anaerobico, mentre oggi si è verificato che questi batteri possano sopravvivere in ambienti ricchi di ossigeno, sono quindi aerotolleranti.
Essi producono energia mediante respirazione anaerobica riducendo i solfati, o altri composti dello zolfo, a idrogeno solforato (H2S).
Il problema relativo a questo gruppo di batteri riguarda importanti aspetti dell’acquariologia e dell’allevamento delle koi in laghetti ornamentali. La presenza in questi ambienti di zone completamente anossiche, concomitante all’assenza di substrati azotati (usati da questi batteri come accettori di elettroni), conduce alla necessità per il batterio di utilizzare composti dello zolfo.
L’idrogeno solforato (o acido solfidrico) è un composto incolore tossico e maleodorante. È una sostanza altamente tossica per i pesci e le piante. Nei pesci la sua azione si esplica a livello cellulare inibendo la respirazione mitocondriale e, legandosi alla emoglobina, impedisce l’accesso dell’ossigeno nel metabolismo cellulare.
L’effetto tossico sulle piante invece non è acuto, ma cronico per la sottrazione di microelementi essenziali per il funzionamento dei sistemi enzimatici.
Batteri Anaerobi Facoltativi : quelli responsabili della denitrificazione dissimilativa (Pseudomonas, Micrococcus, Archromobacter, Bacillus, Alcaligenes)
Sono batteri anaerobi che possono usare l’ossigeno se presente. Il loro metabolismo principale rimane quello anaerobico.
COME TRARRE VANTAGGI DAL CICLO DELL’AZOTO
Il ciclo dell’azoto coinvolge e permea le molteplici reazioni chimiche della natura. È in grado di eliminare, trasformandoli, i prodotti di scarto degli animali.
È quindi naturale pensare che il principio possa essere sfruttato dall’uomo per molteplici funzioni:
Allevamento di pesci d’acqua dolce in laghetti ornamentali e acquariologia
Sin dai tempi antichi l’uomo ha compreso il valore alimentare del patrimonio ittico. Inizialmente, i pesci vennero allevati in cattività per ragioni pratiche, cioè come fonte di cibo.
L’acquariologia nacque quando vennero selezionati alcuni individui, per la loro colorazione o per la loro forma. I primi esempi di questo processo li ritroviamo in Cina, nel VI secolo a.C dove vennero selezionate alcune varietà dorate di carpe, dotate anche di una forma particolare.
In Cina e Giappone l’allevamento non era limitato solo allo scopo alimentare, quanto anche a quello ornamentale in vasche e laghetti da giardino.
L’allevamento acquariologico del pesce rosso (Carassius auratus) era già noto in Cina sotto la dinastia Sung tra 960 e il 1279 D.C. In Europa il suo allevamento si diffonde dopo il 1600, e, in epoca Vittoriana, diventa molto di moda.
Il primo a riconoscere l’importanza dei valori chimici dell’acqua nell’allevamento dei pesci fu Mr. Humphreys, che predisse inoltre l’invenzione del termostato e del riscaldatore. Inoltre introdusse il principio di sovraffollamento, affermando che non era possibile allevare creature viventi in un “pollice quadrato per ognuna”.
Mantenere dei pesci in un sistema di acqua di ricircolo porta alla inevitabile evidenza che gli scarti prodotti dai pesci finiscano per accumularsi diventando tossici per i pesci. Da qui la necessità di riprodurre, in un sistema chiuso, il meccanismo naturale di detossificazione da azoto noto con il nome di ciclo dell’azoto.
Per la trattazione dell’argomento specifico sui filtri rimando a questo articolo: Il variegato mondo dei filtri per laghetto. Qui basta sapere che il procedimento usato sfrutta la nitrificazione come principale via di depurazione.
Tuttavia la trasformazione da ammoniaca a nitrito e poi nitrato produce l’inevitabile conseguenza di una accumulo di quest’ultimo, per cui spesso è necessario l’ausilio di un fitodepurante che attui una denitrificazione assimilativa capace di estrarre l’azoto dall’acqua in virtù di una sua incorporazione nei tessuti della pianta.
Un particolare tipo di filtrazione per laghetto riguarda il filtro anossico. Questo filtro utilizza un substrato argilloso posto in cesti preforati usati per la piantumazione nei laghetti, al cui centro viene posto un “nocciolo” di laterite (rimando agli articoli specifici per una più dettagliata relazione: Filtro Anossico). Questa configurazione è in grado di selezionare naturalmente una specifica tipologia di batteri, quelli anaerobi facoltativi, che sono in grado di effettuare una completa dentitrificazione a partire dai substrati inorganici presenti nel laghetto, l’ammonio e il nitrato.
Questa metodica di filtrazione attua una completa estrazione della molecola dell’azoto, sotto forma di gas N2, rendendo non necessario l’intervento delle piante quale sua fonte assimilativa.
Acquacoltura e ittiocoltura
L’acquacoltura, o più specificamente ittiocoltura, consiste nell’allevamento a scopo riproduttivo di organismi acquatici, principalmente pesci, ma anche crostacei e molluschi, e alghe, in ambienti confinati e controllati dall’uomo. Quando questi allevamenti sono posti all’interno di bacini naturali sfruttano il ciclo di depurazione naturale dell’intero bacino, mentre quando si tratta di allevamenti in vasche isolate è necessario dotarle di filtri che sfruttino le capacità detossificanti dei batteri nitrificanti.
Acquaponica
L’acquaponica è la coltivazione simbiontica di piante e animali acquatici in un ambiente di ricircolo. In pratica consiste nell’allestire una vasca nella quale verranno posti i pesci di allevamento oppure ornamentali, ed impostato un recircolo dell’acqua che passi forzatamente attraverso un’altra vasca in cui venga riposta argilla espansa fino a riempimento. Successivamente in questa vengono posti a dimora ortaggi vari.
Il principio qui sfruttato è la denitrificazione assimilativa attraverso il riassorbimento dei composti azotati dalle radici delle piante, ma anche la nitrificazione, in quanto nell’argilla finiscono per insediarsi e riprodursi i tipici batteri della nitrificazione che attuano un rapporto di mutuo scambio con le radici delle piante. Rimando ad un articolo di approfondimento su questa tematica: Acquaponica, la fitodepurazione utile
Trattamento delle acque reflue
Si definisce trattamento delle acque reflue il processo di rimozione dei contaminanti da un’acqua reflua di origine urbana o industriale, ovvero di un effluente che è stato contaminato da inquinanti organici e/o inorganici.
La depurazione biologica delle acque di scarico consiste nell’azione combinata di diverse specie microbiche che, attraverso vari stadi, degradano le sostanze contenute in esse, trasformandoli in parte in composti semplici come acqua, anidride carbonica, azoto e metano, ed in parte in nuovi organismi.
I primi ritornano in natura, gli altri vengono separati per sedimentazione ed allontanati come fango.Tutti i sistemi di rimozione biologica dell’azoto comprendono una zona aerobica nella quale avvengono le reazioni di nitrificazione. Deve inoltre essere presente una zona anossica nella quale rendere possibili le reazioni di denitrificazione. In questo modo si ottiene la completa rimozione dei composti azotati dal refluo per trasformazione ultima di questi in azoto gassoso.
Il differente posizionamento della vasca anossica distingue i diversi procedimenti biologici gli uni dagli altri.
Diversi sono gli approcci metodologici alla depurazione delle acque, e vengono qui brevemente elencati:
• MBR
• ANAMMOX
• SISTEMI DI AERAZIONE
• FOTODEPURAZIONE
SBR (Sequencing Batch Reactor)
Gli impianti SBR sono dei sistemi di trattamento biologico delle acque a flusso discontinuo dove, all’interno di un unico reattore, avvengono in alternanza le varie fasi del processo comune a tutti i tipi di impianti a fanghi attivi, e non richiede perciò alcun tipo di ricircolo.
Questo tipo di depuratore orienta quindi nel tempo, piuttosto che nello spazio, la depurazione dei reflui, cioè la depurazione si svolge in sequenza temporale anzichè spaziale.
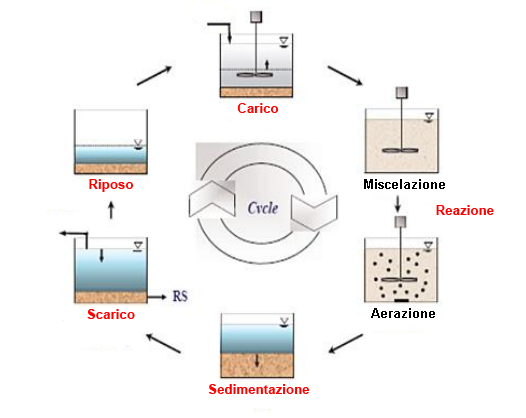 Il maggior pregio dei reattori SBR è quello di poter sfruttare al meglio quello che invece è per i reattori di tipo continuo un problema: la variabilità del carico in ingresso.
Il maggior pregio dei reattori SBR è quello di poter sfruttare al meglio quello che invece è per i reattori di tipo continuo un problema: la variabilità del carico in ingresso.
In fase di avviamento è possibile selezionare una biomassa adattata a diverse condizioni operative alternando periodi di “carestia” e “abbondanza”.
È proprio questa alternanza che che stimola la selezione di batteri capaci di eseguire in modo più efficace le fasi di nitrificazione/denitrificazione.
Le varie fasi della reazione che avvengono nel reattore sono:
MBR (membrane Biological Reactor)
I reattori biologici a membrana rappresentano un sistema ancora relativamente poco sfruttato. Si avvalgono di membrane per la separazione dell’effluente dalla biomassa.
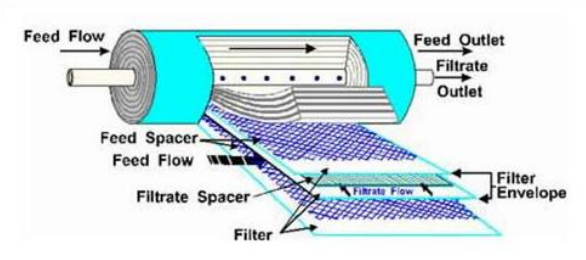 In base alla porosità delle membrane si distinguono due differenti tipologie di filtrazione: Microfiltrazione e Ultrafiltrazione
In base alla porosità delle membrane si distinguono due differenti tipologie di filtrazione: Microfiltrazione e Ultrafiltrazione
La forza che innesca il meccanismo di filtrazione è dato dalla pressione applicata alla membrana tra il lato del liquame e quello del refluo.
Dato che le membrane si sporcano e divengono meno permeabili, è necessario aumentare la tubolenza che diminuisce il ricorso ai controlavaggi delle membrane.
Se la permeabilità diminuisce in modo sensibile allora si interviene con lavaggio con sostanze chimiche.
SISTEMI DI AERAZIONE
L’ossidazione dei fanghi attivi riproduce artificialmente quei processi che si sviluppano naturalmente in un corpo idrico ove vi sia presenza di sostanza organica.
Nei processi a fanghi attivi, pur essendo in gioco gli stessi meccanismi, si evidenzia una sostanziale diversità di ordine quantitativo ripetto ai fenomeni naturali di depurazione. La concentrazione di sostanza organica e di batteri negli impianti raggiunge infatti concentrazioni particolarmente elevate. Ne consegue che, per tenere in vita una tale mole di batteri, divenga necessario rendere disponibile una quantità di ossigeno adeguata e ciò si realizza ricorrendo a sistemi artificiali di aerazione.
L’ossigenazione agisce inoltre generando dei fiocchi prodotti dallo sviluppo della popolazione microbica che sono facilmente sedimentabili.
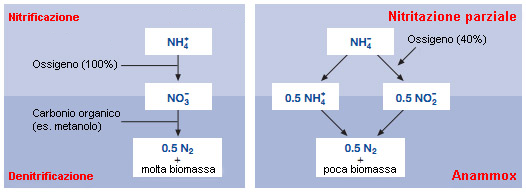
ANAMMOX
La denitrazione anammox viene artificialmente prodotta in appositi reattori nei quali vengono ricreate le condizioni ideali allo sviluppo e riproduzione dei batteri.
Nei processi di tipo anammox solo metà dello ione ammonio è ossidato, e non sino a nitrato, ma solo sino allo ione nitrito (metodo Sharon). Successivamente, lo ione nitrito reagisce con lo ione ammonio rimanente sino a formare azoto molecolare.

I batteri anammox furono per la prima volta osservati in un impianto di depurazione nel 1995.
Sono organismi autotrofi che vivono in ambienti a basso potenziale RedOx ed in condizioni di totale mancanza di ossigeno.
Questi batteri hanno un tasso di crescita molto lento, e sono attivi in un intervallo di pH che va da 6,4 a 8,3 ed una temperatura di 20-43°C, con valori ottimali a 37°C.
A questa temperatura i tempi di replicazione sono dell’ordine di 10-14 giorni.
Confrontato con i normali procedimenti nitro-denitro, il processo anammox presenta dei vantaggi:
alta capacità di estrazione di azoto;
riduzione del 90% delle emissioni di anidride carbonica;
i batteri anammox non richiedono fonti di carbonio organico;
la reazione produce un minore massa biologica da dovere successivamente smaltire;
contenuta superficie di esercizio e ridotti costi di gestione.
FITODEPURAZIONE
La fitodepurazione è un sistema di depurazione usato per le acque reflue domestiche, agricole e talvolta industriali.
 Le tecniche di fitodepurazione possono essere classificate in base alla prevalente forma di vita delle piante acquatiche che vi vengono utilizzate:
Le tecniche di fitodepurazione possono essere classificate in base alla prevalente forma di vita delle piante acquatiche che vi vengono utilizzate:
sistemi a microfite (alghe unicellulari);
sistemi con macrofite galleggianti (presenza di piante acquatiche quali Eichhornia crassipes, Lemna minor, Lemna gibba, Lemna oscura, Lemna trisculca);
sistemi con macrofite radicate emergenti (presenza di macrofite quali Phragmites australis, Typha latifolia, Scirpus lacustris, Scirpus robustus, Scirpus validus).
Questi ultimi sistemi possono subire una ulteriore classificazione dipendente dal cammino idraulico delle acque reflue: sistemi a flusso superficiale o libero; sistemi a flusso sub-superficiale o sommerso a sua volta suddivisi in: sistemi a flusso sommerso orizzontale e verticale.
Nei bacini di fitodepurazione l’acqua può diffondere: a flusso orizzontale, nel quale i liquidi vengono fatti scorrere secondo un piano orizzontale che attraversa il media piantumato; a flusso verticale, nel quale il flusso dell’acqua avviene attraverso un sistema di percolazione attraverso il media piantumato.
La depurazione avviene mediante l’azione combinata tra piante e microrganismi che si insediano sia nel substrato sia nei peli degli apparati radicali. I batteri responsabili della trasformazione dell’ammonio in nitrato sono i batteri nitrificanti. Il procedimento della fitodepurazione si avvale anche dell’effetto diretto di assimilazione della pianta sia di ammonio che di nitrati al fine di inglobarli nella sua stessa struttura. I metalli pesanti vengono infine assorbiti quali sostanze necessarie alla sua crescita.
I vantaggi della fitodepurazione sono i seguenti: costi minimi di costruzione e manutenzione rispetto a quelli degli impianti di depurazione tradizionali; assenza di odori e di proliferazione di insetti nei sistemi a flusso sommerso; totale abbattimento della carica patogena; creazione di un’area verde al posto di manufatti in cemento; possibilità di riutilizzo dell’acqua depurata a scopi irrigui; riduzione dei consumi di energia elettrica rispetto ad un depuratore tradizionale; assenza o ridotta necessità di apparecchiature elettromeccaniche; funzionamento depurativo indipendente dall’assenza di energia o sovraccarico idraulico fondamentali per il funzionamento dei processi tradizionali.
Gli svantaggi sono invece rappresentati da: richiesta di maggiori superfici rispetto ai depuratori convenzionali; costi di acquisizione del suolo sul quale costruire l’impianto.
Bibliografia : L. DE CARLI – Ciclo Biologico dell’Azoto – Metabolismo dell’Azoto Inorganico nelle Piante e nei Batteri – Ed. Piccin
-

By: Lio Fornellino
Il Dr. Franco Prati (1967-2012), medico, era conosciuto nel mondo delle Koi come Lio Fornellino. La sua enorme passione lo aveva portato a fondare il Lio Fornellino Koi Blog dove, pubblicando i suoi interessanti articoli, ci spronava a “guardare” la nostra passione con gli occhi vivi dell’intelligenza. E’ stato anche co-fondatore e primo Presidente dell’IKC – Italian Koi Club.
Alla sua morte abbiamo pensato di dare continuazione alla sua opera di divulgazione e i suoi scritti sono stati le fondamenta della Gazzetta delle Koi.
Un abbraccio caro amico, ovunque tu sia ……. -
-
Propedeutica indispensabile per una diagnosi di malattia
Inaspettato pericolo dalle piante del giardino
Ammoniaca: terrificante sostanza nel laghetto
Nitriti: un riconosciuto pericolo per i pesci
Nitrati
Chilodonella
